UN PEU D’HISTOIRE
L’apparition des premiers végétaux sur Terre est une histoire de symbiose plante / micro-organisme. Il est maintenant établi que les premières plantes qui vivaient alors exclusivement dans les océans ont pu coloniser le milieu terrestre grâce à leur association symbiotique avec des champignons. Ces associations ont donné les premières symbioses que l’on appelle mycorhizes (association plante / champignon). Elles datent de plus de 450 millions d’années Strullu (1985) ; Smith & Read (2008).
Mais l’histoire ne s’arrête pas aux mycorhizes. Les bactéries, levures et champignons étaient déjà présents sur la terre ferme lorsque les plantes ont commencé à s’y développer et leur histoire est depuis lors intimement liée. Cette colonisation microbienne est si ancienne et si répandue que de nombreux mécanismes intègrent des micro-organismes dans leur déroulement quotidien - comme la nutrition, le développement ou l’immunité...
1. PARTENAIRES INDISPENSABLES DES PLANTES
En agriculture, les micro-organismes ont longtemps été perçus exclusivement comme des agents pathogènes. Cette vision négative est désormais désuète et l’étude des communautés microbiennes associées aux plantes nous a permis de mettre en évidence de nombreux micro-organismes d’intérêt agronomique. Présents dans la rhizosphère (zone proche des racines) et dans la phyllosphère (parties situées au-dessus du niveau du sol) certains colonisent également l’intérieur de leur hôte, on les appelle alors endosphériques.
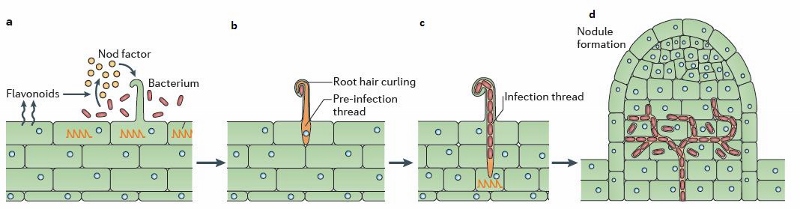
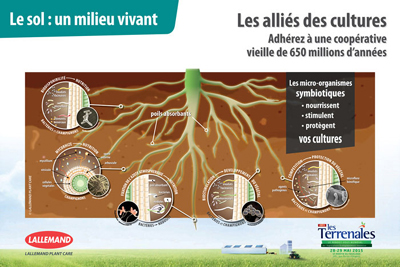
Les micro-organismes rhizosphériques interagissent avec la plante au niveau de ses racines.
C’est là que se trouve la flore microbienne la plus riche et nous aimons faire le parallèle avec notre intestin qui grouille lui aussi d’une flore aussi indispensable que fascinante. Comme un intestin retourné, les racines possèdent des poils absorbants à proximité desquels l’on retrouve d’importantes colonies microbiennes. Ce sont les exsudats racinaires émis spécifiquement par la plante qui attirent et stimulent ces micro-organismes bénéfiques pour elle. Certaines bactéries par exemple, stimulent et/ou protègent la plante grâce à un ou plusieurs mécanismes : excrétion de phytohormones dans le milieu, solubilisation d’éléments minéraux bloqués dans le sol, fixation de l’azote atmosphérique, réduction du niveau de certaines maladies dans le sol (par compétition ou hyper parasitisme…). On appelle ces actéries : PGPR (Plant Growth Promoting Rhizobacteria). C’est dans cet environnement que l’on retrouve également les champignons (mycorhiziens par exemple mais pas seulement) et autres levures...
La phyllosphère et l’endosphère hébergent également une microflore diversifiée impliquée dans des mécanismes de nutrition ou de protection de la plante contre des stress abiotiques et/ou biotiques comme la stimulation des défenses naturelle SDN …
Les micro-organismes ne sont pas des principes actifs ou des fertilisants au sens propre. Ce sont de véritables partenaires des cultures qui tissent avec elles des relations complexes allant bien au-delà de la fonction pour lesquels on les sélectionne. Accompagnée d’une vie microbienne diversifiée, une plante forme à elle seule un véritable écosystème et l’on sait aujourd’hui que plus cet écosystème est complexe, plus il a de chance de résister aux aléas extérieurs.
2. ELOGE DE LA DIVERSITE
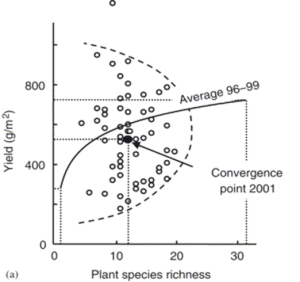
L’étude suivante nous donne un début d’éclairage sur l’importance d’une telle diversité. Bien que stable en apparence, on sait qu’un écosystème ne cesse d’évoluer et qu’il est capable de s’adapter aux modifications de son biotope. Des recherches récentes sur l’évolution du biotope sur les rives du lac miroir (Ceillac) dans les Alpes nous en donnent un excellent exemple avec une analyse remontant au-delà de la dernière glaciation. Cette étude montre comment, contre toute attente, des plantes supérieures ont pu traverser des changements climatiques extraordinairement rudes - l’auteur parle d’une oasis de vie maintenue dans l’espace et le temps – grâce à l’interdépendance et aux interactions de l’immense variété d’organismes vivants dans ce biotope (Leigh Van Valen (1973) et Ehrlich et Raven (1964)). Si cette résilience nous éclaire sur l’importance de la diversité, nous n’en comprenons probablement qu’une faible part aujourd’hui.
3. ET L’AGRICULTURE DANS TOUT ÇA
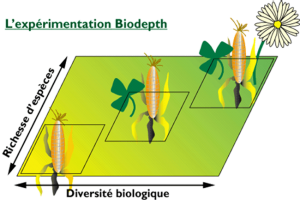
Figure 3L’agriculture entraine une baisse de diversité des micro-organismes due à la simplification et aux variations rapide de la couverture végétale. Phénomène amplifié par l’introduction de plantes non endémiques.
En modelant le paysage et créant de nouveaux biotopes sur des territoires très importants, l’agriculture a bouleversé l’ensemble des équilibres préexistants. Ceci, de façon quasi instantanée à l’échelle du temps nécessaire à la mise en place d’un écosystème stable (minimum 50 ans selon d’IDDR) et sans comparaison avec le temps nécessaire à la mise en place d’une symbiose ou d’un mutualisme par coévolution qui demande lui, plusieurs milliers d’années ! Dans ce contexte, il est peu probable que les plantes agricoles aient une chance de recréer naturellement un écosystème microbien optimal.
L’idée « clef de voûte » cultivée par Lallemand Plant Care est de rechercher puis de proposer des micro-organismes d’intérêt pour les cultures. Grâce à des partenariats scientifiques dans le monde entier, nous étudions les mécanismes sous-jacents aux interactions avec les micro-organismes et sélectionnons par criblage des souches d’intérêt agronomique. Notre objectif est de « compléter » certaines fonctions de la plante par son association avec des micro-organismes clefs dans le domaine de la nutrition et de la résistance aux stress abiotiques et biotiques. Ceci nous conduit à sélectionner, produire, formuler tester et homologuer des micro-organismes permettant d’inoculer artificiellement les cultures.
En associant deux types de génétiques au champ nous pensons pouvoir créer un nouvel organisme hybride (plante / micro-organisme) dont nous espérons un effet d’hétérosis !
Au-delà des inoculations possibles, nous considérons que les rotations ou successions de plantes au niveau parcellaire doivent être prises en compte en tant qu’écosystème variable à l’échelle annuelle ou pluriannuelle mais cohérent sur un temps plus long et c’est dans cette perspective que nous imaginons une action durable et rentable de nos associations plantes / micro-organismes. C’est l’un des points qui différencie fondamentalement l’usage de micro-organismes en agriculture de l’usage des intrants traditionnels.
Cette démarche ne prend toute sa valeur économique et technique que si elle est associée à un raisonnement de la fertilisation (minérale et organique), de la rotation du travail du sol … afin d’exprimer au mieux les potentiels génétiques de la plante et du/des micro-organismes associés.